Cell重磅:破解30年难题,耶鲁团队揭示压力引起IL-6升高对机体功能影响的具体机制
 应激引起炎症反应的利与弊
应激引起炎症反应的利与弊
研究人员通过试管束缚模型、社会隔离模型等急性应激后小鼠血液中的IL-6水平显著升高。并进一步对应激后小鼠血液中的32种炎症细胞因子和趋化因子进行筛选,其中趋化因子CXCL1、细胞因子IL-5、IL-6、GCSF的水平明显增加,但是IL-6是变化最大、也是最显著的细胞因子。
研究人员进一步发现经历过应激的人类组织样本中IL-6也明显增加,但是应激后人类的IL-6水平比应激后小鼠要低100多倍,表明了物种间的差异性。
IL-6由包括造血细胞,心肌细胞,内皮细胞和脂肪细胞等多种细胞类型产生。为了研究急性应激后IL-6的具体来源,研究人员对中脑及前脑等大脑组织,肺、肝脏、心脏、肾脏等外周器官,肌肉、棕色脂肪组织;附睾白色脂肪组织;腹膜后白色脂肪组织等16种组织进行筛选。
结果发现应激小鼠只有棕色脂肪组织IL-6水平出现明显的升高。通过外科手术切除棕色脂肪组织后应激后小鼠的IL-6水平并不升高。这些结果表明应激后外周增加的IL-6主要来自于棕色脂肪组织。
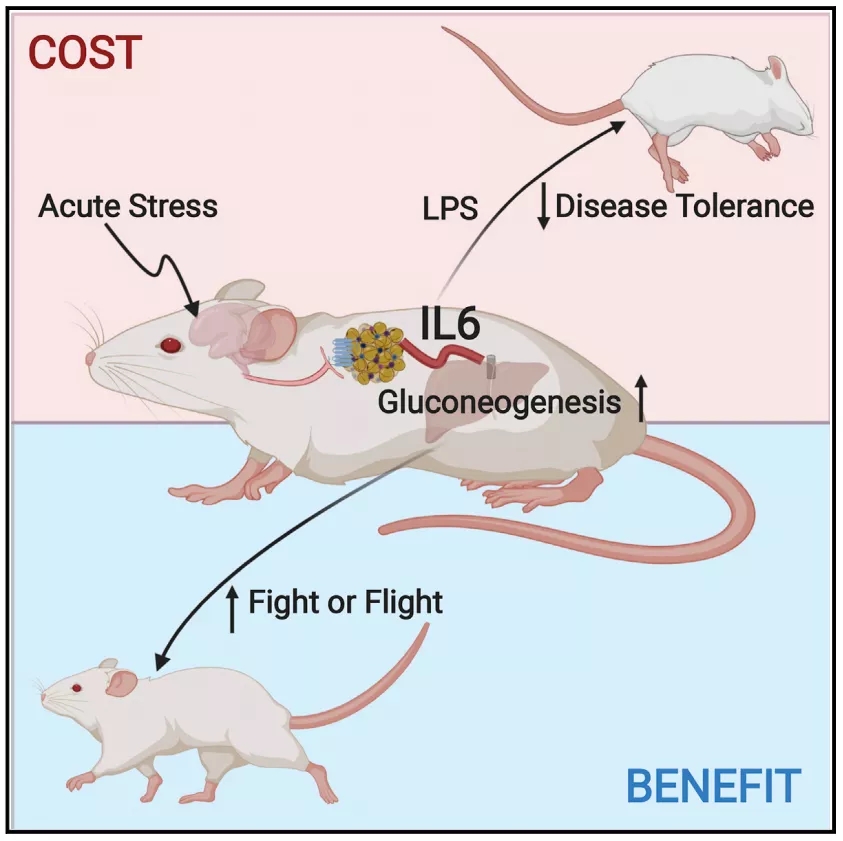
棕色脂肪组织富集表达肾上腺素能受体,肾上腺素能受体通过解偶联蛋白1的帮助下产生热量。研究人员直接分别向小鼠注射肾上腺素能受体1,2,3的激动剂,结果只有肾上腺素能受体3的激动剂能够促进小鼠IL-6水平的升高。此外在应激前给与肾上腺素能受体3的拮抗剂或特异性敲除棕色脂肪组织上的肾上腺素能受体3后小鼠在应激下后IL-6的水平并不会升高,这表明棕色脂肪组织上的肾上腺素能受体3调控应激后IL-6水平的升高。
战斗或逃跑反应(Fight-or-flight response)最早是由美国心理学家怀特•坎农在1929年提出这个概念的,在面对威胁时,产生的一系列应激行为和生理反应。
尽管先前研究表明作为高血糖可以诱导产生IL-6,而血糖增高是急性应激后典型的特征。但是在这项研究中研究人员发现高血糖并不负责IL-6的产生。但是这并不妨碍IL-6通过高血糖调控战斗或逃跑反应。
在面对威胁做出战斗或逃跑反应时需要大量的能量储备,其中葡萄糖最为关键。急性应激后4小时小鼠仍处于高血糖水平,给与IL-6重组蛋白后小鼠产生类似于应激引起的高血糖。在抑制IL-6信号通路后应激小鼠产生葡萄糖的水平降低。
这又带来了一个新的问题:这些葡萄糖从哪里来的?肝脏和肾脏是主要的葡萄糖生产器官,研究人员通过转录组学发现在急性应激过程中只有肝脏诱导葡萄糖产生的基因发生明显变化。此外,这些应激小鼠在接受抗IL-6的受体IL-6Ra抗体处理后,也只有肝脏产生葡萄糖减少。
为了进一步证实在应激后IL-6的升高对于肝脏产生高水平的血糖是必需的,他们特异性敲除肝脏上的IL-6后,在多个应激模型中也观察不到葡萄糖水平的增加,表明了IL-6对肝脏葡萄糖合成的控制作用。
那么损害肝脏的葡萄糖产生是否会影响急性应激的适应性行为?研究人员利用明暗箱实验发现急性应激小鼠呆在黑暗环境中的时间增加,这是一种正常的逃避行为;而在特异性敲除肝脏上的IL-6小鼠并不出现这种增加,这表明应激诱导的IL-6通过IL-6Ra信号调控肝介导应激引起的高血糖,这种正向的能量平衡对于适应性行为反应是必需的。
那么在病理状态下应激产生的IL-6水平升高会加剧炎症反应?研究人员利用LPS(脂多糖)诱导小鼠出现炎症反应,正常情况下这种模型并不会引起小鼠死亡,但是经过试管束缚、社交隔离等急性应激后小鼠死亡率明显增加。更令人意外的是,给与肾上腺素能受体3的激动剂或单剂量IL-6后经过LPS模型的小鼠死亡率也明显增加,这表明IL-6参与其中。而在给与肾上腺素能受体3的拮抗剂后能够明显降低试管束缚引起的高死亡率。也就说明了应激诱导的IL-6降低了宿主对随后的炎症反应的适应性。
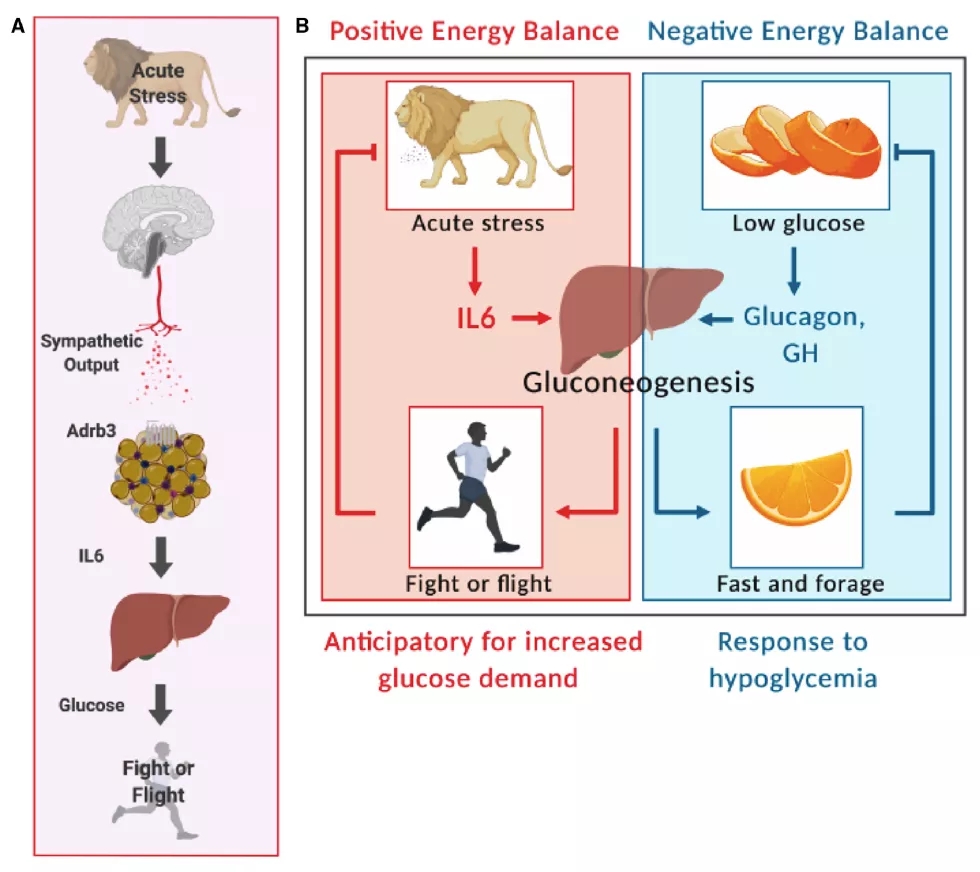
总的来说,本文通过细致的研究阐述了应激诱导的IL-6对机体功能的机制:急性应激引起交感神经输出,激活肾上腺素受体3后促进IL-6水平的升高,随后增强肝脏合成葡萄糖的能力,为逃避或战斗储备能量。
论文链接:https://doi.org/10.1016/j.cell.2020.05.054
2/2 首页 上一页 1 2